Endokrynol. Ped. 13/2014;2(47):57-64
DOI: 10.18544/EP-01.13.02.1486
Płeć i autoimmunizacyjne choroby tarczycy
Klinika Pediatrii i Endokrynologii, Warszawski Uniwersytet Medyczny
Słowa kluczowe: autoimmunizacyjne choroby tarczycy, chromosomy płciowe, hormony płciowe
Streszczenie
Autoimmunizacyjne choroby tarczycy (AIChT) występują znacznie częściej u kobiet niż u mężczyzn, co sugeruje, że dla ich rozwoju istotne znaczenie mają czynniki związane z płcią. Ustalono, że istnieje wyraźny dymorfizm płciowy układu immunologicznego. U kobiet stwierdza się wyższą liczbę limfocytów CD4+, zwiększony stosunek subpopulacji CD4+ do CD8+ i preferencję odpowiedzi zależnej od Th2. U mężczyzn z kolei stwierdza się wyższą aktywność komórek NK oraz preferencję odpowiedzi zależnej od Th1. Podobne zjawiska zaobserwowano na zwierzęcych modelach doświadczalnych. Istnieje wiele hipotez, które mają wyjaśnić potencjalne przyczyny zwiększonej podatności płci żeńskiej na AIChT. Wśród nich za najistotniejsze uważa się czynniki genetyczne związane z chromosomami płciowymi. Na chromosomie X są zlokalizowane geny ważne dla regulacji odpowiedzi immunologicznej: m.in. Foxp3 (Xp11.23), CD40L (Xq26), BTK (Xq21.33-q22), IL2RG (Xq13.1), XIAP (Xq25). Sugeruje się także znaczenie ukierunkowanej inaktywacji drugiego chromosomu X, która występuje częściej u kobiet z autoimmunizacyjnymi chorobami tarczycy. Kolejnym możliwym czynnikiem jest zjawisko mikrochimeryzmu płodowego, który stwierdza się częściej u kobiet z AIChT. Znaczenie może mieć również zaburzenie funkcji microRNA (miRNA), które jest istotnym regulatorem odpowiedzi immunologicznej, a chromosom X zawiera aż 10% całego miRNA genomowego. Wydaje się także, że chromosom Y może pełnić rolę protekcyjną
Wstęp
Zwiększona predyspozycja płci żeńskiej do autoimmunizacyjnych chorób tarczycy jest powszechnie znanym zjawiskiem [1]. Według danych amerykańskich opublikowanych przez National Institutes of Health Autoimmune Diseases Coordinating Committee około 5% populacji krajów zachodnich rozwija choroby autoimmunizacyjne, a 80% tych chorych to kobiety (www.niaid.nih.gov). Autoimmunizacyjne zapalenie tarczycy z niedoczynnością występuje u kobiet z częstością 350/100 000/ rok, u mężczyzn częstość ta wynosi 80/100 000/ rok, natomiast choroba Gravesa- Basedowa jest rozpoznawana z częstością 80/100 000/rok u kobiet i 8/100 000/rok u mężczyzn [2, 3]. Znacząca różnica w częstości występowania tych chorób u płci żeńskiej i męskiej jest widoczna w każdej grupie wiekowej, także w populacji dziecięcej [4], ale dotychczas nie udało się w pełni wyjaśnić jej przyczyn. Istnieje kilka teorii próbujących uzasadnić to zjawisko. Jedną z nich jest bezpośredni wpływ hormonów płciowych na układ immunologiczny.
Zróżnicowanie płciowe układu immunologicznego
Na podstawie licznych badań udowodniono, że istnieje dymorfizm płciowy układu immunologicznego. U płci żeńskiej stwierdza się wyższą liczbę limfocytów CD4+ oraz zwiększony stosunek komórek CD4+ do CD8+ [5]. Makrofagi u kobiet wykazują zwiększoną zdolność do fagocytozy, ponadto mają zwiększoną ekspresję receptorów typu Toll: TLR-2,-3,-4,(TLR, ang. Toll -like receptor), które są odpowiedzialne za inicjację i rozwój odpowiedzi zapalnej [6]. Płeć żeńska wykazuje także preferencję do odpowiedzi immunologicznej humoralnej, zależnej od komórek Th2 i związanej z nasiloną produkcją przeciwciał [7]. U mężczyzn natomiast istotnie wyższa jest aktywność komórek NK i stwierdza się wyraźną preferencję do odpowiedzi komórkowej, zależnej od komórek Th1, z przewagą reakcji cytotoksycznej [7]. Obserwacje te potwierdzono na zwierzęcych modelach doświadczalnych, opartych głównie na myszach wyselekcjonowanych genetycznie. Zatem u osobników żeńskich wykazano żywszą odpowiedź na immunizację, a także zwiększoną produkcję przeciwciał w odpowiedzi na stymulację antygenem. [8]
Wpływ sterydów płciowych na układ immunologiczny
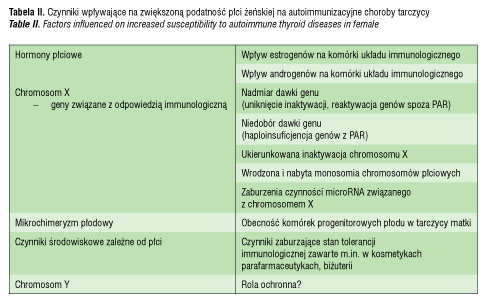
Wydaje się że opisany dymorfizm płciowy układu immunologicznego może być związany z bezpośrednim wpływem hormonów płciowych na komórki immunokompetentne. Udowodniono, że istnieje wzajemna zależność między sterydami płciowymi i limfocytami, a receptory estrogenowe i androgenowe są obecne w wielu komórkach układu immunologicznego. Receptory androgenowe stwierdzono w limfocytach B i T oraz w makrofagach [9]. Z kolei receptory estrogenowe są obecne w limfocytach B i T, komórkach dendrytycznych, neutrofilach, makrofagach, komórkach NK, komórkach macierzy grasicy, komórkach szpiku kostnego, a także w komórkach endotelialnych [7, 10, 11]. Obydwa znane rodzaje receptorów estrogenowych: alfa i beta wykazują zróżnicowaną ekspresję w różnych komórkach układu odpornościowego [7]. Dodatkowo estrogeny wywołują efekt regulacyjny nie tylko na drodze klasycznego receptora w jądrze komórkowym, ale także na drodze błonowego receptora, zależnego od transdukcji sygnału poprzez fosforylację poszczególnych elementów ścieżki sygnałowej, wiodącej przez MAPK [12, 13]. Wszystkie dojrzałe limfocyty mają ekspresję receptorów estrogenowych związanych z błoną komórkową, a limfocyty aktywowane antygenem wykazują zwiększoną transdukcję sygnału z tego receptora [13]. Działanie estrogenów na komórki układu immunologicznego jest wielokierunkowe. Wpływają one na proces dojrzewania limfocytów B i T. Ekspozycja na estrogeny zmniejsza liczbę powstających w grasicy komórek T CD4+ i promuje ich pozagrasiczą limfopoezę. Zaburza to proces kontrolowanej negatywnej selekcji autoreaktywnych limfocytów T i sprzyja ich przetrwaniu, a w konsekwencji wiąże się z ryzykiem przerwania stanu autotolerancji [5].
Estrogeny stymulują produkcję przeciwciał i autoprzeciwciał przez komórki B. Na dowód tego u kobiet stwierdza się wyższe niż u mężczyzn poziomy zarówno IgG, jak i IgM. Efekt ten zaczyna być widoczny po rozpoczęciu dojrzewania płciowego i zanika po menopauzie [14], co sugeruje bezpośredni wpływ hormonów płciowych. Estrogeny stymulują także proliferację komórek T. Osobniki żeńskie w odpowiedzi na infekcję zwiększają liczbę limfocytów T CD4+, w tym Treg CD4+Foxp3+, a indukcja tego procesu jest zależna od sygnału z TLR [15, 16]. Pod wpływem estrogenów zwiększa się ekspresja perforyny w Treg i produkcja IL-10 oraz TGF-beta [16]. Estrogeny hamują odpowiedź komórkową zależną od Th1, ponadto są szczególnie ważne jako modulatory procesu zapalnego. Powodują one hamowanie odpowiedzi indukowanej przez antygeny lipopolisacharydowe i wzmacniają pobudzenie receptora TLR- 4 [7] .
Znacznie mniej wiadomo na temat wpływu androgenów na układ odpornościowy. Jedną z przyczyn trudności w ocenie wpływu androgenów na komórki układu immunologicznego jest fakt, że testosteron, dzięki konwersji przez aromatazę, może wywierać działanie poprzez receptory androgenowe jak i estrogenowe. Stwierdzono, że androgeny stymulują odpowiedź Th1 zarówno u zwierząt doświadczalnych, jak i u ludzi [8]. Pod ich wpływem dochodzi do zwiększenia produkcji IL-2 i zwiększenia aktywacji limfocytów T CD8+ [8].
Działanie estrogenów i androgenów na komórki układu odpornościowego nie wyjaśnia jednak zwiększającej się z wiekiem częstości występowania AIChT ani przewagi zachorowań u płci żeńskiej obserwowanej przed okresem dojrzewania oraz po menopauzie, a więc w okresach kiedy wpływ hormonów płciowych jest znikomy. Obserwacje te sugerują, że w patogenezie zwiększonej podatności płci żeńskiej na AIChT mogą brać udział nie tylko hormony płciowe, ale także czynniki genetyczne, zależne od chromosomów płciowych. Takie założenie wydaje się słuszne, jeżeli weźmiemy pod uwagę, że na chromosomach płciowych X i Y są zlokalizowane geny ważne dla czynności układu odpornościowego i odpowiedzi immunologicznej.
Potencjalne mechanizmy genetyczne podatności płci żeńskiej na AIChT
Istnieje kilka hipotez dotyczących roli chromosomów płciowych w zróżnicowaniu odpowiedzi immunologicznej u kobiet i mężczyzn. Jedną z nich jest wpływ efektu dawki genów obecnych na chromosomie X związanych z odpowiedzią immunologiczną. Są to między innymi takie geny, jak gen receptora IL-2 (IL2RG), gen dla ligandu CD40 (CD40L), gen FoxP3 (forkhead box P3) [17]. Zaangażowania tych genów w odpowiedź immunologiczną dowodzi fakt, że ich uszkodzenie powoduje związane z płcią zaburzenia układu odpornościowego (tab. I). Ponadto obserwacje te potwierdzają istotne znaczenie dawki tych genów dla czynności układu immunologicznego.
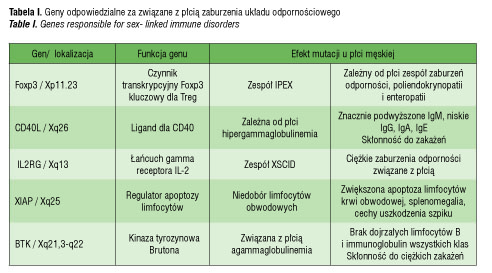
Efekt dawki może polegać na nadmiernej lub niedostatecznej ekspresji poszczególnych genów położonych na chromosomie X. Do nadmiernej dawki może prowadzić uniknięcie inaktywacji przez geny spoza rejonu pseudoautosomalnego chromosomu X lub reaktywacja wyciszonych potranslacyjnie genów w tym rejonie. Natomiast efekt niedostatecznej dawki genu jest związany z utratą genów z rejonu pseudoautosomalnego, co jest określane mianem haploinsuficjencji.
U płci żeńskiej we wczesnym okresie rozwoju płodowego jeden z chromosomów X ulega wyciszeniu potranskrypcyjnemu, czyli inaktywacji. Zjawisko to ma służyć zrównoważeniu dawki genów pomiędzy osobnikami 46, XX i 46, XY. Jedynie rejony pseudoautosomalne PAR1 i PAR2 (ang. pseudoautosomal region) unikają inaktywacji i podlegają rekombinacji w trakcie mejozy, podobnie jak chromosomy autosomalne. Oczekuje się, że podczas randomizowanej inaktywacji dochodzi do zrównoważonego wyciszenia materiału genetycznego z każdego chromosomu X w stosunku 50% do 50%. Proces ten jest randomizowany i jednokrotnego wyboru dla danej linii komórkowej [18]. W wyniku randomizowanej inaktywacji u osobników płci żeńskiej powstaje swego rodzaju mozaicyzm komórkowy, kiedy połowa komórek posiada aktywny chromosom X odmatczyny, a pozostałe komórki aktywny chromosom X odojcowski. W związku z tym w przypadku mutacji jakiegoś genu położonego na chromosomie X u osobnika męskiego będzie ona obecna we wszystkich komórkach, natomiast u płci żeńskiej powinna występować tylko w połowie komórek.
Jedną z genetycznych teorii, które tłumaczą zwiększoną predyspozycję do AIChT u kobiet, jest zjawisko ukierunkowanej inaktywacji drugiego chromosomu X (ang. skewed X chromosome inactivation). Polega ono na nierandomizowanym wyciszeniu jednego z chromosomów X w stosunku innym niż 50% /50%. Skrajnie ukierunkowana inaktywacja może osiągać nawet stosunek przekraczający 90% /10%. W takiej sytuacji istnieje ryzyko, że inne antygeny będą prezentowane w grasicy, a inne w komórkach obwodowych układu odpornościowego, czego skutkiem może być nieprawidłowa identyfikacja antygenów własnych i obcych, a w konsekwencji rozwój procesu autoimmunizacyjnego. Przy skrajnie ukierunkowanej inaktywacji X niemal wszystkie limfocyty T występujące na obwodzie zawierają dane pochodzące z chromosomu X od jednego rodzica i odczytują antygeny pochodzące od drugiego rodzica jako obce. Potwierdzeniem tej hipotezy jest częstsze występowanie ukierunkowanej inaktywacji u kobiet z chorobami autoimmunizacyjnymi tarczycy [19–22], a także u żeńskich par bliźniąt z AIChT [23]. Według metaanalizy Simmondsa i wsp. [24] ukierunkowana inaktywacja drugiego chromosomu X u pacjentek z chorobą Gravesa oraz z chorobą Hashimoto występuje istotnie statystycznie częściej, a iloraz szans jest w obu chorobach zbliżony i wynosi odpowiednio OR=2.5 i OR=2.4 [24]. Dodatkowo stwierdzono, że częstość występowania ukierunkowanej inaktywacji chromosomu X zwiększa się z wiekiem, co mogłoby dodatkowo tłumaczyć zjawisko zwiększającej się z wiekiem częstości autoimmunizacyjnych chorób tarczycy [25].
Kolejną hipotezą, która może wyjaśnić szczególną predyspozycję płci żeńskiej do AIChT, jest zjawisko monosomii chromosomu X. Najlepszym przykładem wrodzonej monosomii X są pacjentki z zespołem Turnera, u których obserwuje się zwiększone ryzyko autoimmunizacyjnych chorób tarczycy [26]. Opisywana jest także nabyta mononosomia chromosomu X w wybranych liniach komórkowych. Invernizzi i wsp. [27] stwierdzili, że odsetek obwodowych komórek krwi z monosomią chromosomu X jest istotnie wyższy u pacjentek z AIChT w porównaniu do zdrowych kobiet. Monosomia X jest obserwowana częściej w obwodowych limfocytach T i B aniżeli w innych komórkach krwi, co zwiększa ryzyko zaburzeń autotolerancji. W wyniku nabytej monosomii geny zależne od X unikające inaktywacji wykazują haploinsuficjencję genów z rejonu PAR. Dane te sugerują, że zjawisko haploinsuficjencji genów zależnych od chromosomu X może być głównym czynnikiem warunkującym zwiększoną częstość występowania chorób autoimmunizacyjnych u kobiet [27].
Z kolei efekt nadmiernej dawki genów jest widoczny u osobników z aberracjami chromosomów płciowych związanymi z obecnością dodatkowych chromosomów. Przykładem są pacjenci z zespołem Klinefeltera (47, XXY), u których ryzyko chorób autoimmunizacyjnych jest zbliżone do ryzyka u płci żeńskiej [28]. Ryzyko to jest szczególnie wysokie dla tocznia układowego. Pacjenci z zespołem Klinefeltera wykazują zmniejszoną liczbę komórek NK w porównaniu do mężczyzn 46,XY, co wydaje się bezpośrednio związane z obecnością dodatkowego chromosomu X [28].
Z powstawaniem efektu dawki może mieć związek działanie microRNA (miRNA) obecnego na chromosomie X. Szacuje się, że chromosom ten zawiera aż 10% całego miRNA genomowego, podczas gdy chromosom Y jest całkowicie pozbawiony miRNA [29]. Można przypuszczać że zaburzenia dotyczące miRNA mogą wpływać na efekty działania genów związanych z odpowiedzią immunologiczną, położonych na chromosomie X, które ulegają wyciszeniu po transkrypcji [29]. MicroRNA jest to małe dwuniciowe, niekodujące RNA biorące udział w regulacji postranskrypcyjnej. Ogranicza translację, przez co jest silnym regulatorem odpowiedzi immunologicznej. Reguluje odpowiedź pierwotną na patogeny przez osłabienie sygnału z receptorów błonowych typu Toll (ang. Toll-like receptors, TLRs), co z kolei ogranicza niespecyficzną odpowiedź zapalną. Zaburzenia ekspresji miRNA mogą modyfikować odpowiedź na antygeny zarówno w odpowiedzi immunologicznej pierwotnej, jak też adaptacyjnej [30]. Znaczenie miRNA w patogenezie AIChT u płci żeńskiej wydaje się tym bardziej istotne, że istnieje wzajemne oddziaływanie pomiędzy estrogenami, miRNA i receptorami TLR. Estrogeny regulują ekspresję miRNA, promując odpowiedź zapalną w splenocytach poprzez receptory typu alfa i wpływają na czynność TLR [31]. Przypuszcza się też, że miRNA indukowane przez estrogeny wpływa na siłę stymulacji ich receptora i hamuje odpowiedź na stymulację estrogenami przez negatywne sprzężenie zwrotne [30, 31]. Te wzajemne zależności tworzą sprzyjające warunki do rozwoju procesu autoimmunizacyjnego u płci żeńskiej.
Kolejna koncepcja próbująca wyjaśnić szczególną predyspozycję płci żeńskiej do AIChT zakłada wpływ zjawiska mikrochimeryzmu płodowego, czyli przemieszczania się komórek płodu do krążenia matczynego podczas ciąży, w tym macierzystych komórek szpiku i komórek układu immunologicznego. Te płodowe komórki progenitorowe przedostają się do tkanek matczynych i mogą indukować odpowiedź immunologiczną, co prowadzi do rozwoju procesu autoimmunizacyjnego. Potwierdzeniem tej hipotezy jest fakt, że u kobiet z chorobą Hashimoto i chorobą Gravesa- Basedowa istotnie częściej stwierdza się w tkance tarczycowej obecność mikrochimeryzmu płodowego [32–34].
W ostatniej dekadzie w analizie czynników predysponujących płeć żeńską do chorób autoimmunizacyjnych bierze się pod uwagę nie tylko te, które zwiększają ryzyko, ale także możliwość ochronnego działania materiału genetycznego zawartego na chromosomie Y. Geny obecne w rejonie pseudoautosomalnym, PAR1 i PAR2 chromosomu Y są homologiczne do ich odpowiedników na chromosomie X i mają istotne znaczenie dla prawidłowej czynności układu immunologicznego. Są to między innymi: gen dla receptora alfa IL-3, (IL3RA) konieczny do prawidłowego rozwoju komórek progenitorowych szpiku, gen dla receptora IL-9 (IL9R) obecnego na limfocytach Th2 i Th17 oraz Treg. Persani i wsp. [35] uważają, że u chorych płci męskiej z AIChT może dochodzić do utraty rejonu pseudoautosomalnego chromosomu Y, co powoduje haploinsuficjencję genów z PAR1 i PAR2 w analogicznym mechanizmie, jak w przypadku monosomii chromosomu X u płci żeńskiej. W swoim badaniu stwierdzili, że u mężczyzn z AIChT częściej niż w zdrowej grupie kontrolnej dochodzi do utraty chromosomu Y w limfocytach obwodowych [35]. Można więc przypuszczać, że wraz z utratą chromosomu Y są oni pozbawieni czynnika, który chroni płeć męską przed chorobami autoimmunizacyjnymi. Te obserwacje wymagają jednak dalszych wnikliwych badań, gdyż postępująca z wiekiem nabyta haploinsuficjencja genów zależnych od chromosomów płciowych w komórkach układu immunologicznego może być objawem starzenia się układu immunologicznego [36]. Proces ten mógłby tłumaczyć rosnącą z wiekiem podatność na infekcje, nowotworzenie i choroby autoimmunizacyjne u obu płci.
Czynniki środowiskowe zależne od płci
Wydaje się, że na zwiększone ryzyko rozwoju chorób autoimmunizacyjnych, w tym AIChT, u kobiet mają wpływ nie tylko czynniki hormonalne, genetyczne, epigenetyczne, ale także środowiskowe, które są zależne od płci (tab. II). Mogą one wynikać z typowego dla danej płci zawodu, który stwarza ryzyko ekspozycji na czynniki potencjalnie modulujące układ odpornościowy. Do immunomodulatorów należą także związki zawarte w kosmetykach i parafarmaceutykach, a także metale [37], np. miedż i złoto, z których wytwarza się biżuterię. Metale mogą zmieniać odpowiedź immunologiczną zarówno przez bezpośredni wpływ na limfocyty, jak też pośrednio przez indukowanie ekspresji nowych antygenów i modyfikację kompleksów prezentujących antygen [38].
Podsumowanie
Ustalenie czynników wpływających na zwiększoną podatność płci żeńskiej na choroby autoimmunizacyjne, a także czynników które prawdopodobnie mają ochronny wpływ na zmniejszone ryzyko tych chorób u mężczyzn potencjalnie otwiera nowe możliwości terapeutyczne. Próby wykorzystania terapii hormonami płciowymi do korzystnego modyfikowania przebiegu choroby były już podejmowane w chorobach autoimmunizacyjnych, które prowadzą do znacznego stopnia niesprawności i trwałego uszkodzenia narządów. Estrogeny i leki modyfikujące receptory estrogenowe są zalecane w niektórych postaciach reumatoidalnego zapalenia stawów. Korzystny wpływ zastosowania estrogenów stwierdzono także w eksperymentalnym autoimmunizacyjnym zapaleniu mózgu u genetycznie wyselekcjonowanych myszy z ryzykiem stwardnienia rozsianego [39]. Szczególną rolę przypisuje się estriolowi, głównemu estrogenowi ciążowemu, który może generować tolerogenne komórki dendrytyczne zapobiegające rozwojowi procesu autoimmunizacyjnego [40]. Z kolei doświadczalne autoimmunizacyjne zapalenie tarczycy u myszy ulega zaostrzeniu po podaniu estrogenów, a testosteron ma łagodzący wpływ na dynamikę przebiegu choroby [39]
Być może w przyszłości wiedza na temat związanych z płcią czynników predysponujących do chorób autoimmunizacyjnych, w tym także chorób tarczycy, pozwoli na opracowanie skutecznych zindywidualizowanych metod leczenia, które będą uwzględniały zarówno rodzaj schorzenia, jak i płeć pacjenta.
Piśmiennictwo
1. Gleicher N., Barad D.H.: Gender as risk factor for autoimmune diseases. J. Autoimmun., 2007 Feb:28(1), 1- 6
2. Jacobson D.L., Gange S.J., Rose N.R. et al.: Epidemiology and estimated population burden of selected autoimmune diseases in the United States. Clin. Immunol. Immunopathol., 1997 Sep:84(3), 223-243.
3. Cooper G.S., Stroehla B.C.: The epidemiology of autoimmune diseases. Autoimmun. Rev., 2003 May 2 (3), 119-123.